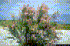 |
|
|
 |
|
|

|
|
|
Tamarix ramosissima (tree, shrub) |
|
|
Taxonomic name: Tamarix ramosissima (Ledeb.)
Synonyms: Tamarix pallasii var. brachystachys Bunge, Tamarix pentandra
Common names: salt cedar (English), Sommertamariske (German), tamarisk (English), tamarix
Organism type: tree, shrub Tamarix ramosissima is a rampantly invasive shrub that has dominated riparian zones of arid climates. A massive invasion of T. ramosissmia in the western United States has dominated over a million acres. Typically found in conjunction with other Tamarix species and resultant hybrids, T. ramosissima displaces native plants, drastically alters habitat and food webs for animals, depletes water sources, increases erosion, flood damage, soil salinity, and fire potential. Description
Tamarix ramosissima is a semi-deciduous, loosely branched shrub or small to medium-sized tree. The branchlets are slender with minute, appressed scaly leaves. The leaves are rhombic to ovate, sharply pointed to gradually tapering, and 0.5 - 3.0mm long. The margins of the leaves are thin, dry and membranaceous. Flowers are whitish or pinkish and borne on slender racemes 2-5cm long on the current year's branches and are grouped together in terminal panicles. The pedicels are short. The flowers are most abundant between April and August, but may be found any time of the year. Petals are usually retained on the fruit. The seeds are borne in a lance-ovoid capsule 3-4mm long; the seeds are about 0.45mm long and 0.17mm wide and have unicellular hairs about 2mm long at the apical end. The seeds have no endosperm and weigh about 0.00001 gram. (Carpenter, 2003; Dudley, pers. comm.).T. ramosissima, Tamarix aralensis, and T. chinensis can be distinguished from other members of Tamarix by their sessile leaves, pentamerous flowers, and hololophic androecial discs. T. chinensis and T. ramosissima can be distinguished from T. aralensis by its caducous petals at the time of seed maturation.T. ramosissima and T. chinensis may be distinguished by a few microscopic floral characters especially where the filament is inserted into the nectary disk and edaphic affinities. T. ramosissima has an eroded denticulate, obovate petals, and is halophilous, while T. chinensis has entire sepals, elliptic-ovate petals, and prefers non-halophilous soils (Gaskin & Scheel, 2003) Similar Species
Tamarix aphylla, Tamarix canariensis, Tamarix chinensis, Tamarix gallica, Tamarix parviflora
More
Occurs in:
agricultural areas, coastland, desert, estuarine habitats, lakes, riparian zones, ruderal/disturbed, urban areas, water courses, wetlands General impacts
Tamarix ramosissima has displaced or replaced native plant communities and may be a major contributor to the decline of many native plants and animals, including endangered species (Dudley & Deloach, 2004). Alteration of natural flooding regimes through dam construction has resulted in T. ramossisima replacing many native tree species, such as cottonwood (Populus deltoides subsp. wislizenii) and willows (Salix spp.), in riparian forests (Everitt 1980; Horton 1977; Robinson 1965; Graf 1978). The invasion of Tamarix ramosissima along streams is likely to have altered the food webs in these aquatic ecosystems (Kennedy & Hobbie 2004). The roots of T. ramosissima bind together gravel and cobble riverbeds, resulting in enlarged bars and narrowed channels increasing the likelihood of flood (Cooper et. al 2003). The leaf litter and foliage produced by T. ramosissima is flammable and encourages the spread of wildfires (Busch 1995; Brotherson & Field 1987; Dudley et al 2000). Native vegetation and wildlife is destroyed in these fires, while T. ramosissima seedlings are able to increase their spread. This is due to their ability to re-sprout more successfully than native plants following fire (Huntert et al 1988; Busch 1995; Ellis 2001; Dudley et al 2000). T. ramosissima is capable of utilizing saline groundwater by excreting excess salts through glands in the leaves causing an increase in surface soil salinity. This increase, combined with dense canopy of saltcedar plants and higher likelihood of fires within stands of saltcedar, results in the elimination of native riparian plants (APHIS, 2000).
T. ramosissima is also known to transpire large amounts of groundwater, which dessicates soils and reduces the water table. Its transpiration rate is similar to native plants on a per-leaf basis but it maintains a larger leaf area per ground area, and therefore uses more water in total (Sala et al 1996; Dahm et al 2002; Shafroth et al 2005; Cleverly et al 2002). Because T. ramosissima can take up water from non-saturated soils, it has an added advantage in outcompeting native vegetaion (Dudley, pers. comm.).
T. ramosissima posseses many physiological adaptations that allow it to replace the native tree species, especially along human-altered river stretches. These include: high seed production, rapid germination and seedling establishment, high growth rates, high ET rates, drought tolerance, extreme salt tolerance, flood tolerance, the ability to resprout after fire, and high leaf area index (LAI) allowing it to establish quickly and deplete water-tables at the expense of native species These advantages appear to be so overwhelming that, once it becomes established, eradication of it by human intervention is difficult but necessary to restore riparian corridors (Glen & Nagler, 2005). Uses
Often planted as an ornamental and to prevent erosion in arid areas. Tamarix ramosissima provides a nectar source for honeybees in some areas, and is widely used in the old world for furniture making and for firewood, for tannin extraction, and for cover for livestock (Dudley, pers. comm.). T. ramossisima may also be useful for bioremediation, for instance it takes up perchlorate from groundwater, perchlorate being a pollutant derived from jet fuel (Urbansky et al. 2000).Many species of native birds, including the endangered and federally protected south-western willow flycatcher (Empidonax traillii extimus), are able to exploit T. ramosissima for shelter and nesting, especially when some native trees remain (Fleishman et al. 2003). However, it is mostly foliage gleaners and fairly opportunistic species that use it to a substantial extent - cavity nesters like owls and wrens, drillers like woodpeckers and sapsuckers, frugivores, granivores and other specialists rarely occupy tamarisk (Ellis 1995, Shafroth et al. 2005, Hunter 1984, Hunter et al. 1985, Cohan et al. 1979, Lovich and DeGouvenain 1998, Dudley and DeLoach 2005) and usage by insectivores declines greatly as vegetation dominance by tamarisk increases (Yard et al. 2004). Notes
There are few plants that are true genetic species of Tamarix ramosissima in infested areas, at least in North America. Most of what is called T. ramosissima represents a variety of hybrids, including haplotypes of T. ramosissima, T. chinensis, T. gallica and others (Gaskin and Schaal 2002); it even hybridizes with athel (T. aphylla), an evergreen species, in some southwest U.S. locations (Gaskin and Shafroth, in press). The most common genotype in the U.S. is a morphologically cryptic hybrid of T. ramosissima and T. chinensis not detected in Eurasia (Gaskin & Schaal, 2002). Geographical range
Native range: Temperate Asia: Afghanistan, Iran, Iraq; Caucasus: Armenia, Azerbaijan, Russian Federation -Ciscaucasia, Dagestan, Kazakhstan, Kyrgyzstan, Tajikistan, Turkmenistan, Uzbekistan, Mongolia; China: China - Gansu, Nei Monggol, Ningxia, Qinghai, Xinjiang, Xizang. Tropical Asia: Pakistan; East Europe: Moldova; Russian Federation - European part; Ukraine (USDA ARS, 2009).
Known introduced range: Naturalised in South Africa, United States (USDA ARS 2009); Reported in parts of Canada (Natureserve, 2009); Mexico (CONABIO 2008); Argentina (Dewine & Cooper, 2008); Australia (Department of the Environment and Heritage and the CRC for Australian Weed Management, 2003). Introduction pathways to new locations
For ornamental purposes: Introduced as ornamentals and for windbreaks (Sobhian et. al 1998).
Landscape/fauna "improvement": Introduced as ornamentals and for windbreaks (Sobhian et. al 1998).
Nursery trade: Introduced as ornamentals and for windbreaks (Sobhian et. al 1998).
Local dispersal methods
On animals (local): Massive quantities of minute seeds are readily dispersed by wind (Carpenter 2003).
Other (local): Tamarix ramosissima's growth rate substantially increased where beavers (Castor canadensis) had reduced the canopy of native plants in eastern Montana (Kimball & Perry, 2008). Similar concerns were expressed in Grand Canyon National Park, Arizona (Mortenson et al 2008).
Translocation of machinery/equipment (local): Tamarix ramosissima seeds and other plant propagules may be transported by earth-moving equipment during site construction between and the and by boats and their towing vehicles (Pearce & Smith 2003). Management information
Mechanical: Hand pulling can be used where plants are small, access is difficult, or herbicides cannot be used (Carpenter 2003). Uprooting methods are effective in the short-term because uprooted trees do not resprout. For sawing and mowing, chemical treatment may be necessary to prevent resprouting. Immature plants may often be physically removed by hand with care given to complete removal of the root structure and disposal of the plant by burning or deep burial. Bulldozing, followed by root-plowing is successful, consistent and effective when used on large thickets of established Tamarix ramosissima. Managed flooding can effectively kill T. ramosissima on a long-term basis. Repeated flooding is necessary to kill saltcedar seedlings that are rapidly established from windborne seeds. Established saltcedar plants can tolerate flooding for up to 3 months. Conditions suitable for controlled flooding exist in relatively small areas such as highly managed wildlife refuges (APHIS, 2003). Chemical: Aerial application of the herbicide imazapyr, alone or in combination with glyphosate, is effective and practical for controlling T. ramosissima over thousands of hectares, particularly in dense stands where little or no native vegetation is present. Several field trials have produced control rates of > 90% after one or two years (Carpenter 2003). On smaller sites the cut stump method is successful when triclopyr herbicides are also used. Basal bark applications of Garlon4 were very effective on plants with a basal diameter of less than 4 inches. Burning, followed by herbicide application to the resprouts, also produced excellent results, although this method is not appropriate when T. ramosissima exists as a component of native plant communities (Carpenter 2003). The use of triclopyr (Garlon4 or Remedy) mixed with oil and applied as a basal bark or cut stump treatment has been used with great success on scattered infestations, with no resprouting occurring. The basal bark treatment involves applying the herbicide mixture to the lower 18 inches of the plant clear to the ground. Herbicides used at aquatic sites include Arsenal and Habitat. These are very effective as foliar treatments, but are not selective and must be used with care. Around 30% of tamarisk may resprout after three years when using these herbicides (Baker, 2005. pers. comm.). Biological: Cattle (and probably goats) will eat T. ramosissima.
A biocontrol agent, the saltcedar leaf beetle (Diorhabda elongate), has been released in nine states (California, Oregon, Nevada, Utah, Wyoming, Colorado, Montana, New Mexico and Texas), excluding those areas where the endangered southwestern willow flycatcher (Empidonax traillii extimus is nesting in tamarisk (Dudley et al. 2001, DeLoach et al. 2004). The Athel Pine National Best Practice Management Manual brings together the best management practices available to date on control options for athel pine (T. aphylla), tamarisk (T. ramosissima) and smallflower tamarisk (T. parviflora). It also illustrates successful control programs with case studies that demonstrate how these weeds are managed effectively in Australia. Included are pointers to identify the Tamarix species you are dealing with as each of them are managed using different strategies. The manual includes a 'Decision Support Tree for Tamarix control' to develop a control program for athel pine, tamarisk or smallflower tamarisk based on the type of infestation you have to treat and the options available to you. Nutrition
Tamarix ramosissima is a facultative phreatophyte, meaning that its roots are able to reach deep water tables but it is capable of tolerating periods without access to water (Carpenter 2003). Reproduction
Tamarix ramosissima is highly fecund. It produces massive quantities of minute seeds that are readily dispersed by wind (Carpenter 2003) but are usually only viable for a few days (Dudley pers. comm.). T. ramosissima seeds have no dormancy or after-ripening requirements. Germination can occur almost immediately upon reaching a moist site, and germination conditions are broad, good germination being found from 10 to 35�C, but mid-summer seed collections indicated poorer germination rates than those collected in late spring (Young et al. 2004). T. ramosissima flowered in two flushes, one in April-May and another in late July in northern Arizona, presumably reflecting availability of spring snowmelt and summer monsoon moisture. This species flowered continuously under favourable environmental conditions but the flowers require insect pollination to set seed (Carpenter 2003). Lifecycle stages
Tamarix ramosissima will produce roots from buried or submerged stems or stem fragments. This allows the species to produce new plants vegetatively following floods from stems torn from the parent plants and buried by sediment. Ideal conditions for first-year survival are saturated soil during the first few weeks of life, a high water table, and open sunny ground with little competition from other plants. The seedlings of this species grow more slowly than many native riparian plant species and it is highly susceptible to shading (Carpenter, 2003). This species has been nominated as among 100 of the "World's Worst" invaders Reviewed by: Tom Dudley Marine Science Institute University of California Santa Barbara & Natural Resource & Environmental Sciences University of Nevada, Reno. United States Principal sources: Carpenter, 2003 Element Stewardship Abstract for Tamarix ramosissima Ledebour
Compiled by: Profile revision: National Biological Information Infrastructure (NBII) & IUCN/SSC Invasive Species Specialist Group (ISSG)
Updates with support from the Overseas Territories Environmental Programme (OTEP) project XOT603, a joint project with the Cayman Islands Government - Department of Environment
|
|
Last Modified: Monday, October 04, 2010
|
|
|
|
 |
|